Guest Commentary by Cristian Proistosescu, Peter Huybers and Kyle Armour
tl;dr
Two recent papers help bridge a seeming gap between estimates of climate sensitivity from models and from observations of the global energy budget. Recognizing that equilibrium climate sensitivity cannot be directly observed because Earth’s energy balance is a long way from equilibrium, the studies instead focus on what can be inferred about climate sensitivity from historical trends. Calculating a climate sensitivity from the simulations that is directly comparable with that observed shows both are consistent. Crucial questions remain, however, regarding how climate sensitivity will evolve in the future.
Background
The recent papers, by Kyle Armour (hereafter A17) and by us (Proistosescu and Huybers, 2017) (hereafter PH17), build on a large literature documenting the time-dependence of climate feedbacks in models. They make quantitative apples-to-apples comparisons between the climate sensitivities simulated by CMIP5 models and those inferred from global energy budget observations.
[Aside: For brevity, we refer to the global energy budget constraints as “observations”. In reality, these constraints are not purely observational; they combine surface and ocean temperature observations over the recent transient evolution of the climate system with model-based estimates of historical radiative forcing and of the global energy imbalance during early-industrial climate. Further, inference of climate sensitivity from these observations rely upon a physical model that entails some strong simplifications.]
Because feedbacks may change over time as patterns of warming evolve, observations made today do not necessarily provide estimates of the long-term, equilibrium climate sensitivity (ECS). Rather, they constrain a quantity that we call the inferred (A17), or instantaneous (PH17), climate sensitivity (ICS). The two studies were performed independently using distinct methodologies, and both find that ICS values are systematically lower than ECS values within CMIP5 models. Moreover, they find that model-derived ICS values are consistent with ICS values inferred from observations.
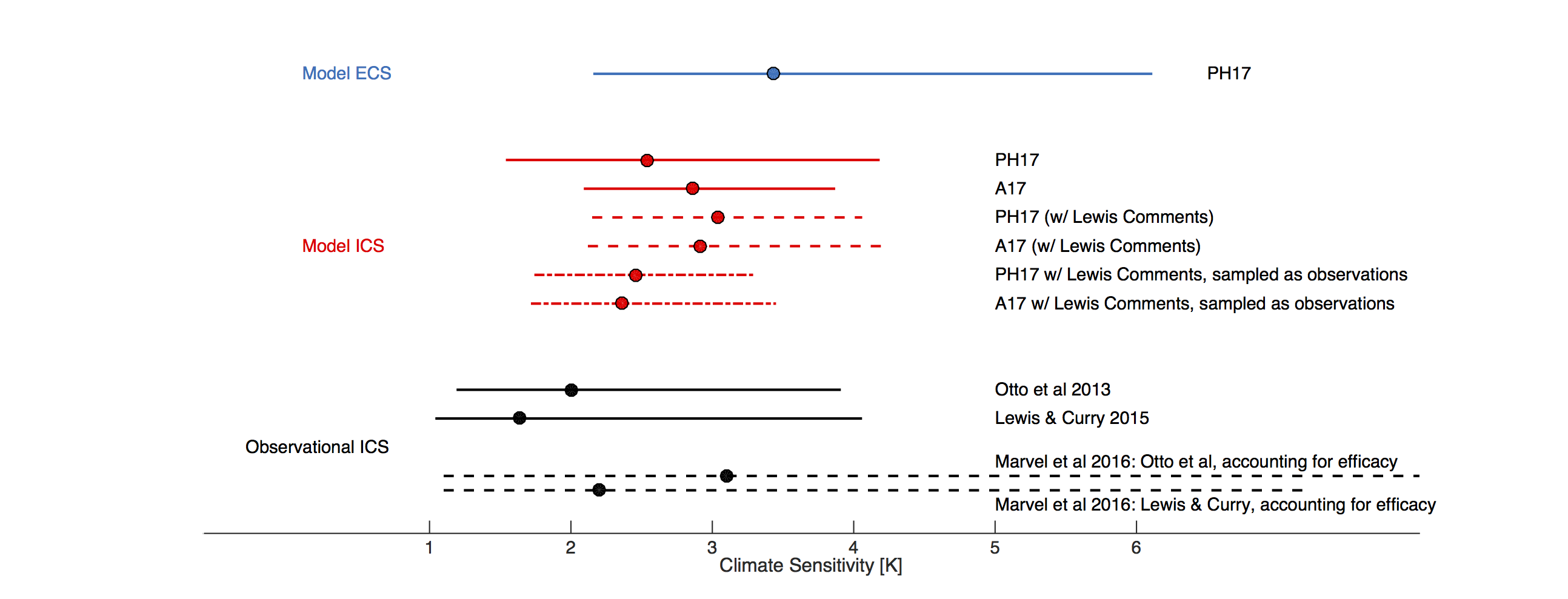
Figure shows model Equilibrium Climate Sensitivity (ECS, blue, from PH17), compared with observationally- and model-derived Inferred/Instantaneous Climate Sensitivity (ICS, black and red). Circles denote medians, while the line denotes the 5-95% confidence interval. Solid lines indicate published estimates, dashed lines indicate PH17 and A17 values with Nic Lewis’ comments taken into account, and dot-dashed lines indicate PH17 and A17 values with both the Lewis correction and the Richardson et al (2015) correction.
Of mountains and molehills
Nic Lewis has posted some criticisms of both papers (A17 critique, PH17 critique) where he proposes changes to these studies that supposedly render the models and data again in disagreement. The criticisms focus on the treatment of radiative forcing within the models which is used in the calculation of ICS values. We approach these criticisms by exploring their implications for the resulting ICS estimates, without necessarily implying agreement.
For A17, Lewis suggests that one should (i) assume that CO2 forcing increases slightly faster than logarithmically with CO2 concentration, rather than linearly, as is traditional[1], and (ii) estimate model CO2 forcing values by a different method[2]. Accounting for these two effects increases model-mean and median ICS by ~0.1°C. For PH17 the suggestions are to (i) adjust historical forcing to account for the average pre-industrial volcanic forcing[3], (ii) rescale the radiative forcing associated with a CO2 doubling for each model when calculating ICS[4], and (iii) better account for the difference between instantaneous and effective radiative forcing, a point that was raised in PH17. While the last suggested adjustment almost certainly conflates time-dependence of feedbacks with stratospheric and tropospheric adjustment, we nonetheless explore the impact of adding Lewis’ upward correction of 0.1°C.
[1]A17 employed the standard assumption that CO2 forcing increases linearly with the logarithm of concentration, while recent work suggests that CO2 forcing might increase slightly faster than linearly. Assuming this forcing nonlinearity decreases model ICS by 0.04°C, on average.
[2]A17 estimated CO2 forcing using linear regression of global radiative imbalance on global temperature over years 1-5 of each model’s abrupt CO2 quadrupling simulations. This choice was made to avoid forcing estimates being biased by the nonlinear relationship between radiative imbalance and temperature that typically emerges beyond year 5. Using years 1-20 for this calculation, as Lewis suggests, seems unwise to us. But doing so increases model ICS by 0.09°C, on average. A third suggestion was to include additional CMIP5 models in the analysis; while we do not do so here, we estimate that including them would bring model-mean ICS back to near the A17 value since most of the additional models are variants of those already included that happen to have low ICS.
[3]Temperature change is considered as an anomaly with respect to pre-industrial climate. Thus, volcanic forcing should also be considered as anomalies with respect to the average pre-industrial forcing. The issues of pre-industrial volcanic forcing can be largely avoided by considering the same averaging interval as in Lewis and Curry (2015), raising model ICS by an average of 0.1°C.
[4]Radiative forcing PH17 inferred from the CMIP5 models (which averages 3.9 W/m2) is larger for a doubling of CO2 than the estimate generally used elsewhere, including in Lewis’s estimates (3.7 W/m2, ref. AR5 TS & 9.7.1, which relies on a subset of the CMIP5 models). In order to make results comparable, PH17 rescaled radiative forcing to the commonly-used AR5 value.
In the figure, we show both the original A17 and PH17 estimates of model ICS, and versions that take into account Lewis’s suggestions. The most notable change is that the PH17 model-median ICS estimate increases from 2.5 to 3.0°C, while the 5-95% credible interval narrows from 1.6 to 4.2°C to a range of 2.2 to 4.0°C. This shift is primarily in response to rescaling radiative forcing using model specific values, and it leads to better agreement between the median values of PH17 and A17. However, regardless of the methodology used, or whether or not Lewis’ suggestions are accounted for, the core results still hold: model values of ICS are consistent with observational ICS values.
Should we be concerned by the remaining offset?
Although consistent, the ranges of historical and model based ICS values do not completely overlap. However, there are additional effects that could be taken into account that would make the estimates more congruent. When sampled in a manner consistent with observations, by accounting for spatial coverage and differences between sea-surface and near-surface-air temperatures, changes in model temperature over the historical period are reduced by a median of 19% (i.e. Richardson et al (2015)). A proportional correction to model ICS is illustrated in the figure. Additionally, there is significant uncertainty in the magnitude and efficacy of non-CO2 forcing agents. There is enough uncertainty in fact, that accounting for the efficacy can broaden the range of observational ICS to easily contain all model estimates (Figure 1).
Furthermore, while model-median ICS is based on the forced response in a model ensemble, the observational ICS depends on a single realization of Earth’s warming, the details of which are sensitive to the phase of natural variability affecting the pattern of surface warming over the periods considered. Thus, the observed estimate of ICS is almost certainly perturbed from its expected value (i.e., what would be obtained from perfect knowledge of an infinite ensemble of Earth histories). Recent evidence suggests that observed temperature patterns in recent decades, associated with a cooling of the East Pacific, can lead to a more negative cloud feedback than that seen in the historical response in models over the same period. It seems important to determine whether the inability of models to simulate these recent patterns is indicative of model deficiencies or simply a consequence of natural variability.
Since both estimates have differing sources of uncertainty, demanding agreement between the best-estimate historical ICS and best-estimate model ICS would constitute over-fitting. Additionally, given the fact that the model ensemble is an ensemble of opportunity, we don’t expect the model range to cover the full uncertainty range. Still, it is worth considering the lack of model coverage in the range of low ICS. This discrepancy may reflect that while low values of ICS are allowed by observational constraints (associated with a low aerosol forcing), it is challenging to construct climate models that produce such low ICS values, as it requires a more-negative cloud feedback that is difficult to reconcile with current mechanistic understandings. Better constraints on historical aerosol forcing and forcing efficacy might go a long way towards understanding the plausibility of low ICS values (e.g. Marvel et al, 2016).
What is the magnitude of ECS? (hint:≠ICS)
An important core finding of A17 and PH17 is that values of ICS drawn from the historical record are not sufficient to constrain values of ECS. Indeed, within models, ICS and ECS differ as the strength of radiative feedbacks change over time as patterns of surface temperature evolve with warming. PH17 demonstrated that portions of the climate system that respond over centennial timescales (such as the southern oceans) are important amplifiers of climate sensitivity in the models – a slow-mode response leading to values of ECS that are higher than the values of ICS that reflect more transient warming. Increasing sensitivity over time seems to be associated with a low-cloud feedback excited by warming in the Eastern Equatorial Pacific and Southern Ocean. This slow-mode response (and thus ECS) is essentially unconstrained by global energy budget observations because warming in these regions has been small, possibly held back by upwelling water from the ocean interior.
Key research targets should be improving understanding of (i) how the east-west temperature gradient in the Pacific Ocean will evolve in the future, and (ii) how low-level cloud (and other) feedbacks will respond, in turn. Zhou et al (2017) suggest that feedbacks can vary with the surface warming pattern, at least on decadal timescales. Yet, it is not known whether the magnitude by which feedbacks evolve within the models is realistic. A major challenge going forward is to develop instrumental observations capable of constraining these processes. Another potential path to better understanding slow-mode contributions to ECS is to explore paleoclimate changes occurring over timescale where the climate has had more time to fully come into equilibrium (e.g., PALAEOSENS).
As usual, it will be the interplay between theoretical exploration and observational analysis that will build our understanding of climate sensitivity. Lewis’ comments have helped to sharpen consideration of the topic, but we need not make mountains of difference out of molehills. Model ICS estimates strongly overlap with observed ICS uncertainty ranges, and proposed alterations involve only small modifications. More interesting at this point is to explore how future warming could diverge from historical patterns, and how climate feedbacks in the future might be different from those at present.
Acknowledgements
We thank Nic Lewis and Gavin Schmidt for comments on a draft of this post. We want to note that on the basis of as-of-yet unpublished research, Nic disagrees with our use of the Kummer and Dessler upward correction of observational ICS due to aerosol efficacy.
References
K.C. Armour, “Energy budget constraints on climate sensitivity in light of inconstant climate feedbacks”, Nature Climate Change, vol. 7, pp. 331-335, 2017. http://dx.doi.org/10.1038/nclimate3278
C. Proistosescu, and P.J. Huybers, “Slow climate mode reconciles historical and model-based estimates of climate sensitivity”, Science Advances, vol. 3, pp. e1602821, 2017. http://dx.doi.org/10.1126/sciadv.1602821
C.A. Senior, and J.F.B. Mitchell, “The time-dependence of climate sensitivity”, Geophysical Research Letters, vol. 27, pp. 2685-2688, 2000. http://dx.doi.org/10.1029/2000gl011373
T. Andrews, J.M. Gregory, and M.J. Webb, “The Dependence of Radiative Forcing and Feedback on Evolving Patterns of Surface Temperature Change in Climate Models”, Journal of Climate, vol. 28, pp. 1630-1648, 2015. http://dx.doi.org/10.1175/jcli-d-14-00545.1
P.M. Forster, “Inference of Climate Sensitivity from Analysis of Earth’s Energy Budget”, Annual Review of Earth and Planetary Sciences, vol. 44, pp. 85-106, 2016. http://dx.doi.org/10.1146/annurev-earth-060614-105156
M. Richardson, K. Cowtan, E. Hawkins, and M.B. Stolpe, “Reconciled climate response estimates from climate models and the energy budget of Earth”, Nature Climate Change, vol. 6, pp. 931-935, 2016. http://dx.doi.org/10.1038/nclimate3066
G. Myhre, E.J. Highwood, K.P. Shine, and F. Stordal, “New estimates of radiative forcing due to well mixed greenhouse gases”, Geophysical Research Letters, vol. 25, pp. 2715-2718, 1998. http://dx.doi.org/10.1029/98gl01908
B. Byrne, and C. Goldblatt, “Radiative forcing at high concentrations of well-mixed greenhouse gases”, Geophysical Research Letters, vol. 41, pp. 152-160, 2014. http://dx.doi.org/10.1002/2013gl058456
N. Lewis, and J.A. Curry, “The implications for climate sensitivity of AR5 forcing and heat uptake estimates”, Climate Dynamics, vol. 45, pp. 1009-1023, 2014. http://dx.doi.org/10.1007/s00382-014-2342-y
J.R. Kummer, and A.E. Dessler, “The impact of forcing efficacy on the equilibrium climate sensitivity”, Geophysical Research Letters, vol. 41, pp. 3565-3568, 2014. http://dx.doi.org/10.1002/2014gl060046
K. Marvel, G.A. Schmidt, R.L. Miller, and L.S. Nazarenko, “Implications for climate sensitivity from the response to individual forcings”, Nature Climate Change, vol. 6, pp. 386-389, 2015. http://dx.doi.org/10.1038/nclimate2888
C. Zhou, M.D. Zelinka, and S.A. Klein, “Impact of decadal cloud variations on the Earth’s energy budget”, Nature Geoscience, vol. 9, pp. 871-874, 2016. http://dx.doi.org/10.1038/ngeo2828
R. Knutti, R. Furrer, C. Tebaldi, J. Cermak, and G.A. Meehl, “Challenges in Combining Projections from Multiple Climate Models”, Journal of Climate, vol. 23, pp. 2739-2758, 2010. http://dx.doi.org/10.1175/2009jcli3361.1
J.R. Kummer, and A.E. Dessler, “The impact of forcing efficacy on the equilibrium climate sensitivity”, Geophysical Research Letters, vol. 41, pp. 3565-3568, 2014. http://dx.doi.org/10.1002/2014GL060046
K.C. Armour, J. Marshall, J.R. Scott, A. Donohoe, and E.R. Newsom, “Southern Ocean warming delayed by circumpolar upwelling and equatorward transport”, Nature Geoscience, vol. 9, pp. 549-554, 2016. http://dx.doi.org/10.1038/ngeo2731
E.J. Rohling, E.J. Rohling, A. Sluijs, H.A. Dijkstra, P. Köhler, R.S.W. van de Wal, A.S. von der Heydt, D.J. Beerling, A. Berger, P.K. Bijl, M. Crucifix, R. DeConto, S.S. Drijfhout, A. Fedorov, G.L. Foster, A. Ganopolski, J. Hansen, B. Hönisch, H. Hooghiemstra, M. Huber, P. Huybers, R. Knutti, D.W. Lea, L.J. Lourens, D. Lunt, V. Masson-Demotte, M. Medina-Elizalde, B. Otto-Bliesner, M. Pagani, H. Pälike, H. Renssen, D.L. Royer, M. Siddall, P. Valdes, J.C. Zachos, and R.E. Zeebe, “Making sense of palaeoclimate sensitivity”, Nature, vol. 491, pp. 683-691, 2012. http://dx.doi.org/10.1038/nature11574
 jQuery(document).ready(function(){
jQuery(document).ready(function(){